Mitochondrial membrane potential peptides research sits at an intersection of bioenergetics and molecular pharmacology that has drawn serious attention from cell biologists over the past two decades. The mitochondrial membrane potential, often abbreviated as ΔΨm, refers to the electrochemical gradient maintained across the inner mitochondrial membrane. This gradient is not incidental. It drives ATP synthesis, regulates calcium signaling, and serves as a real-time indicator of overall mitochondrial health. When researchers began exploring how peptides might influence or preserve this potential, the implications reached across multiple fields, from cellular aging to metabolic dysfunction research.
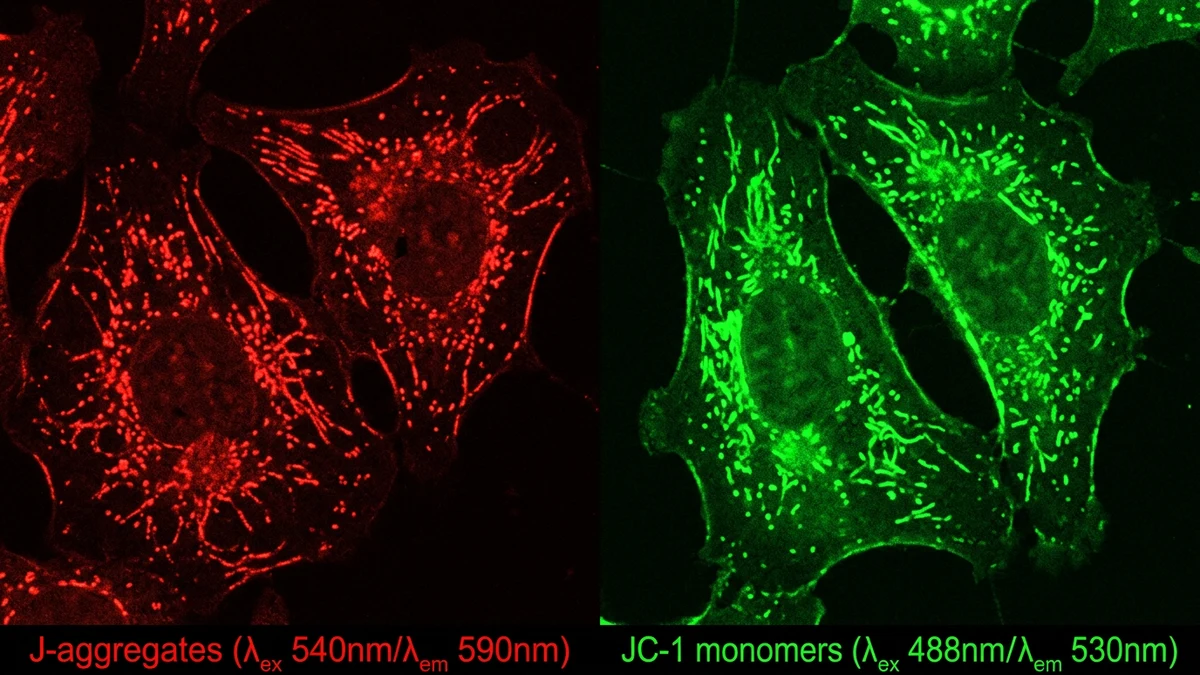
Understanding why this matters requires a brief grounding in what the membrane potential actually represents. Mitochondria generate ΔΨm by pumping protons across the inner membrane through the electron transport chain. This creates a charge difference, negative on the matrix side, that powers the ATP synthase complex. If that potential collapses or fluctuates erratically, the downstream effects touch nearly every energy-dependent process in the cell. Researchers use fluorescent dyes and JC-1 assays to measure ΔΨm in live cells, and shifts in these readings are often among the earliest detectable signs of mitochondrial stress.
ΔΨm is more than a byproduct of respiration. It actively regulates whether a cell undergoes apoptosis, how efficiently it handles reactive oxygen species, and how well it maintains mitochondrial dynamics like fission and fusion. Disrupted membrane potential has been observed in models of neurodegenerative conditions, metabolic syndrome, and ischemia-reperfusion injury, which is why it appears so frequently as a readout in preclinical studies.
Peptide researchers find this particularly relevant because short amino acid chains can cross membranes, accumulate in specific subcellular compartments, and interact with protein complexes in ways that small-molecule drugs sometimes cannot replicate. Some peptides carry mitochondria-targeting sequences derived from naturally occurring signal peptides, allowing them to concentrate at the inner membrane. This targeting specificity is a genuine advantage in research models, even if translating it to clinical applications presents its own challenges.
One acknowledged limitation in this field is that most membrane potential data comes from in vitro systems or rodent models. The relationship between cultured cell findings and whole-organism physiology is rarely straightforward. Researchers working with human subjects face additional complexity because ΔΨm is not easily measured non-invasively, and proxy biomarkers carry their own interpretive uncertainty.
Several categories of peptides have attracted attention in ΔΨm research. SS peptides, sometimes called Szeto-Schiller peptides, were designed specifically to concentrate in the inner mitochondrial membrane through an interaction with cardiolipin, a phospholipid unique to that location. Research suggests these compounds influence electron transport chain function and reduce mitochondrial ROS production in cell culture models, effects that correlate with maintained or restored membrane potential. SS-31, the most studied member of this class, has been examined in animal models of cardiac and renal ischemia, where ΔΨm collapse typically precedes cell death.
Separately, researchers have explored MOTS-c, a peptide encoded within mitochondrial DNA itself. This is unusual. Most peptides studied in a mitochondrial context are exogenous compounds synthesized in laboratories. MOTS-c appears to regulate nuclear gene expression related to metabolism and stress response, and some research suggests it influences how cells maintain mitochondrial homeostasis under conditions of metabolic stress. Its relationship to membrane potential is indirect but reflects how interconnected mitochondrial signaling actually is.
Humanin is another mitochondrially encoded peptide that has appeared in aging and neuroprotection research. Studies in animal models suggest it may support mitochondrial integrity under apoptotic stress, though the exact mechanisms remain an active area of investigation. Both humanin and MOTS-c connect to broader questions about how the mitochondrial genome communicates with the rest of the cell, a topic sometimes called mitochondrial-nuclear crosstalk.
The mechanisms by which peptides influence ΔΨm fall into a few general categories. Some act at the level of the electron transport chain, stabilizing complex activity or reducing electron leak. Others appear to modulate the mitochondrial permeability transition pore, a channel whose opening leads to rapid dissipation of the membrane potential and is closely linked to cell death signaling. A third category interacts with mitochondrial membrane lipid composition, particularly cardiolipin, which plays a structural role in organizing respiratory chain supercomplexes.
Cardiolipin oxidation is a meaningful research target because it destabilizes the cristae architecture where ATP synthesis occurs. When cardiolipin is damaged by peroxidation, respiratory chain efficiency drops and membrane potential fluctuates. SS-31 binds cardiolipin directly, and research suggests this interaction helps preserve cristae structure in stressed cells. The connection between lipid composition and bioenergetic function is an area where peptide research intersects with nutritional science, since dietary fatty acids also influence membrane lipid profiles.
Calcium handling is another mechanism worth understanding here. Mitochondria buffer cytosolic calcium, and this process is driven by the electrochemical gradient of ΔΨm. When membrane potential is high, calcium uptake is efficient. When it drops, calcium dysregulation can follow, triggering cascades that researchers associate with cellular dysfunction. Peptides that support ΔΨm maintenance may therefore influence calcium signaling as a secondary effect, though attributing specific outcomes to single mechanisms in complex biological systems is always interpretively difficult.
Mitochondrial dysfunction is one of the recognized hallmarks of cellular aging, as outlined in landmark reviews by Lopez-Otin and colleagues published in Cell. ΔΨm tends to decline in aged cells across multiple tissue types, and this decline correlates with reduced ATP production capacity and increased mitochondrial ROS generation. The question that peptide researchers are asking is whether exogenous compounds can slow or partially reverse these changes at the cellular level.
This connects naturally to ongoing work in NAD+ precursor research, since the NAD+/NADH ratio directly affects electron transport chain function and therefore membrane potential. Researchers studying peptides that target mitochondria sometimes design experiments alongside NAD+ pathway interventions to understand whether these approaches have additive or independent effects. The interaction between redox state and membrane potential is close enough that separating them experimentally requires careful controls.
Metabolic health research also ties in here. Mitochondria in skeletal muscle are central to glucose metabolism and fatty acid oxidation. Research in animal models of insulin resistance consistently shows impaired ΔΨm in muscle cells, preceding measurable changes in whole-body glucose tolerance. Whether peptide interventions that restore ΔΨm in these models translate to metabolic outcomes remains an open question, one the field has not yet answered with sufficient human data.
Measuring ΔΨm accurately is technically demanding. JC-1 is the most widely used fluorescent probe. It shifts from green to red emission as membrane potential increases, and the ratio of these signals provides a quantitative readout. But JC-1 has known limitations, including sensitivity to light exposure and potential artifacts from probe aggregation. More recent probes like TMRM offer better quantitative precision in live-cell imaging but require careful calibration against known depolarizing agents like CCCP.
Researchers designing peptide studies around ΔΨm outcomes need to account for the fact that many peptides are themselves amphipathic or carry charges that could influence membrane potential as a non-specific effect. Appropriate controls, including scrambled peptide sequences with matched charge profiles, help distinguish specific mechanistic effects from generic membrane interactions.
Delivery is another practical consideration. Many peptides degrade rapidly in biological fluids. The targeting approaches used in SS peptide research, where charge and lipophilicity are tuned to facilitate inner membrane accumulation, represent one solution. Other researchers have explored cell-penetrating peptide carriers or nanoparticle encapsulation to improve mitochondrial delivery. Each approach introduces its own variables that need to be characterized before attributing observed ΔΨm changes to the peptide of interest.
The field is still developing consensus on which experimental models best predict physiologically relevant outcomes. Primary cells behave differently from established cell lines, and animal models vary in how closely their mitochondrial physiology mirrors human tissue. These methodological questions don't undermine the research but do mean that findings require careful contextual interpretation before any broader claims can be supported.
For researchers and practitioners following this space, the most credible signal comes from studies that combine multiple endpoints: ΔΨm measurements alongside ATP quantification, ROS levels, and functional cellular readouts rather than relying on a single biomarker. The convergence of evidence across these independent measures is what separates a genuinely informative result from an artifact of measurement.
Reproducibility across independent laboratories is also worth emphasizing. Several high-profile findings in mitochondrial biology have failed to replicate cleanly, partly because small differences in cell handling, passage number, or probe calibration can shift outcomes meaningfully. The peptide research community has been working to develop more standardized reporting practices, and publications in journals like Redox Biology and Mitochondrion now increasingly require deposition of raw data to facilitate independent verification. This methodological maturation is a positive sign for the field's long-term credibility and utility.
This article is for informational and research purposes only. Nothing here constitutes medical advice, a treatment recommendation, or endorsement of any specific compound or protocol. Peptide research is conducted in regulated laboratory and clinical settings, and findings from preclinical models do not necessarily translate to human applications. Consult qualified healthcare and research professionals before designing or participating in any study involving biological agents. For research purposes only — not medical advice.